研究室紹介
植物遺伝育種学研究室
ゲノム情報を利・活用してブランド特産作物を創り出す

さまざまな生物のゲノム配列の解読が急速に進むようになり、経済的効果が大きい形質(重要形質)に関わる遺伝子の情報がどんどん蓄積されています。最近では、蓄積された重要形質に関わるゲノム情報を活かすことで、新しい植物育種技法が開発されるようになっています。その中には、慣行の交雑育種に要する育種年限の短縮を目的に、ゲノム情報の高度利用によるデータ駆動型の育種や、突然変異育種における変異体作出効率を高める技法があります。前者においては、従来の多大な労力を必要とした表現型選抜にかわり、遺伝型に基づき有用な育種素材を効率的に見出すことが可能となっています。後者においては、ゲノム編集技術が代表的であり、標的遺伝子にピンポイントで変異を導入することにより、優良な重要形質をもつ作物を創りだすことが可能です。 本研究室では、ソバ属植物を研究材料として、農業上有用な形質の探索とその形質が現れるメカニズムを分子レベルで解明し、主に前者の技法を用いて品種改良に役立てようとする研究を進めています。それによって、地域のブランドとなる特産作物の創出を目指しています。
研究概要
フツウソバ(Fagopyrum esculentum)における生殖様式の遺伝的基盤の解明
植物の多くは、受粉時に自己の花粉と非自己の花粉を識別し、自己の花粉との受精を抑制する自他識別システムを有しています。この自他識別システムは自家不和合性とも呼ばれ、その情報ネットワーク制御が如何に行われているのか未解明な部分が多いです。その中でもCharles Darwinの著書「Different forms of flowers」(Darwin 1877)に掲載されているように、花の形態多型と自家不和合性が密接に関連した異形花型自家不和合性が知られています。異形花型自家不和合性は、他の植物でも見られる虫媒による他殖性強化のための巧妙な機構であり、花型と自家不和合性を統御する隣接遺伝子群がS座に集約されて機能していると考えられています(S-supergene; Barrett 2002)。これまでに、私たちは、異形花型自家不和合性を示すフツウソバを材料として、S座に座乗する遺伝子S-ELF3を発見しました(Yasui, Aii ら 2012)。さらに、S-ELF3を失うと異形花型自家不和合性の表現型が、どのように変化するのかを調べるために、世界から収集されたフツウソバ遺伝資源の中から、S-ELF3を欠失した系統を探しました。そして、S-ELF3が欠失し、その周辺領域に変異が生じているS-del1を見出すに至りました。野生型のフツウソバは、雌しべと雄しべの長さが異なる花(短柱花と長柱花)を咲かせ、両者の間でしか受精が成立しません(図1)。ところが、S-del1は、雌しべと雄しべの長さが等しい花(等長柱花)を咲かせ、自殖することができます。さらに、S-del1の花粉は、短柱花と長柱花に対しても受精することができる能力をもちます。最近、Jeffrey ら(2023)によって、S-ELF3は、フツウソバの異形花型自不和合性における雌性側の形質を制御する遺伝子であることが証明されました(図 2)。このことから、S-del1は、S-ELF3を失うことで、等長柱花の花を咲かせ自殖することができるようになったと考えられます。しかし、S-del1の花粉がなぜ短柱花と受精できるのかは不明です。私たちはS-del1を用いたゲノム科学・分子遺伝学的な解析をすることで、フツウソバにおける異形花型自家不和合性の遺伝的基盤の解明を目指しています。
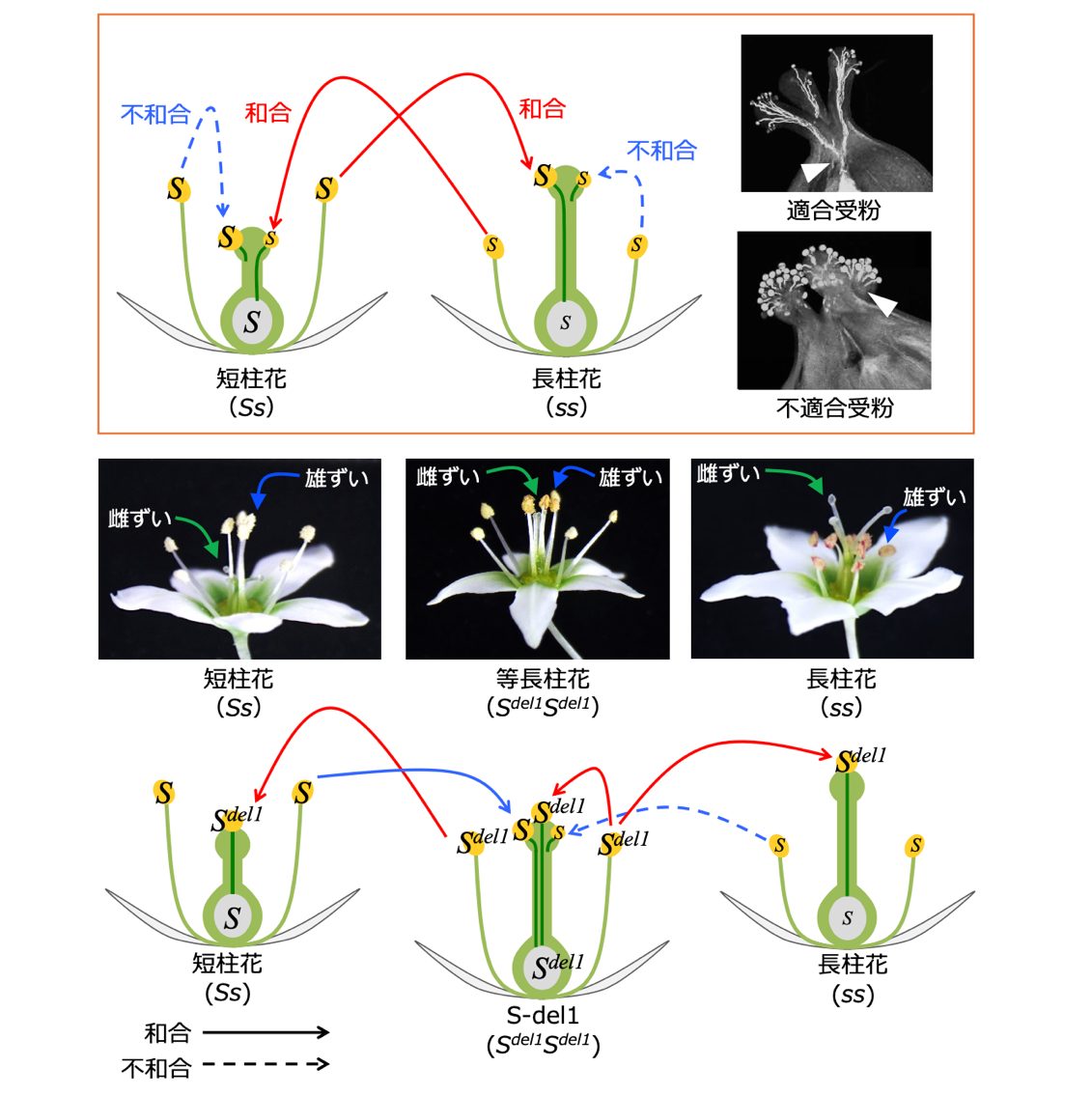
図1 フツウソバにおける異形花型自家不和合性
フツウソバは、雄しべが長く雌しべが短い花(短柱花)と、雄しべが短く雌しべの長い花(長柱花)をもつ個体が存在する異形花型自家不和合性植物です。短柱花のS座の遺伝型はSs、長柱花の遺伝型はssであり、両者間の交配により後代の集団において両花型の存在が1:1に維持されます。私たちが発見した自家和合性変異体S-del-1は、雄しべと雌しべの長さが等しい花(等長柱花)を咲かせます。S-del1のS座の遺伝型は、Sdel1Sdel1です。S-del1の花粉は、短柱花個体及び長柱花個体と受精することが可能です。
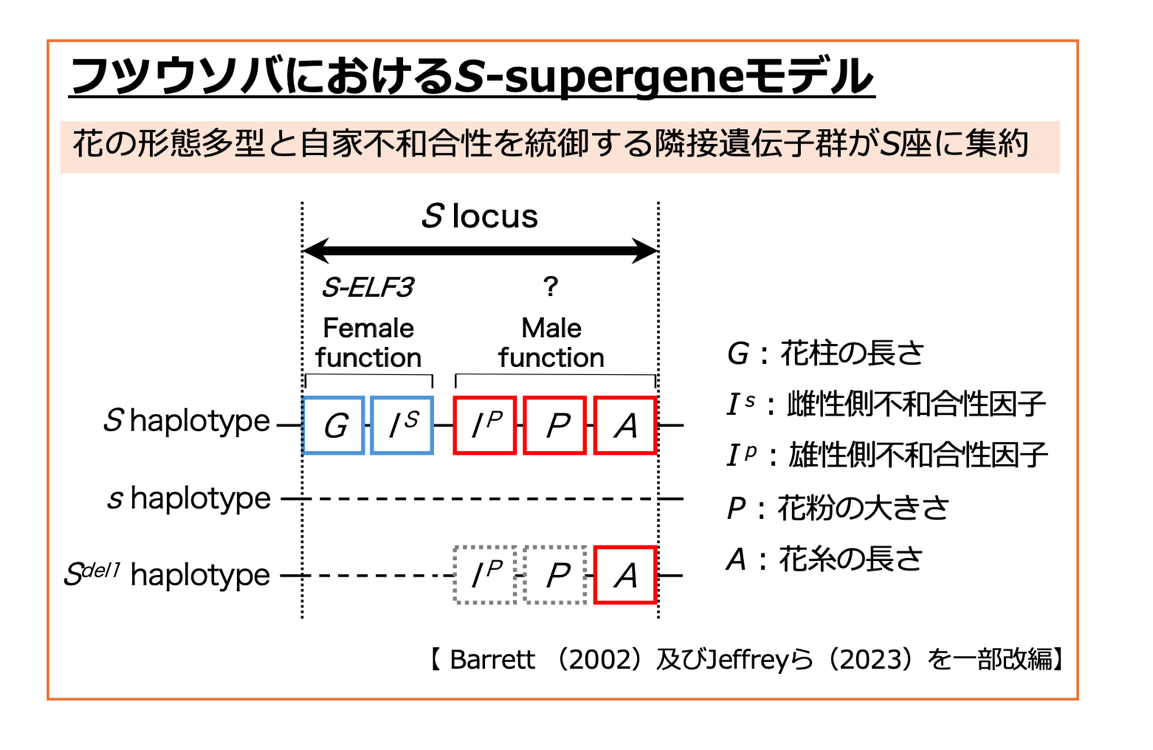
図2 フツウソバにおけるS-supergene
フツウソバのS座は、Sハプロタイプがヘミザイガスの状態であり、sハプロタイプの当該領域は欠失しています。自家和合性変異体S-del1のS座は、S-ELF3が座乗する周辺領域が欠失したSdel1ハプロタイプをもちます。フツウソバS座の雌性側因子は、S-ELF3であることがJeffreyら(2023)によって証明されました。しかしながら、雄性側因子の実体は、未だ明らかになっていません。
気候変動に対応した新形質フツウソバの開発
現時点での世界人口は80億人を突破しており、今後も増加し続け2050年には97億人に達すると予測されています。現在、世界の人間のカロリー需要の三分の二は、三大穀物であるイネ(Oryza sativa)、コムギ(Triticum aestivum)、トウモロコシ(Zea mays)によって供給されています。しかし、今後の地球温暖化の進行が、これら三大穀物の収穫量を著しく低下させる懸念があり、食料安全保障を三大穀物のみに依存していることが問題視されています。この問題の解決は、国連の持続可能な開発目標(SDGs)の中でも、目標2(飢餓を終わらせ、食糧安全保障及び栄養改善を実現し、持続可能な農業を促進する)として挙げられています。このような食料事情から、各地域で古くから栽培はされていますが、品種改良が十分に行われてこなかった作物への関心が高まっています。そこで私たちは、日本の伝統的行事とも関わりが深く、健康志向性食品としても認知されているフツウソバに着目し、食糧安全保障及び栄養改善を実現する品種の創出を試みています。まず、フツウソバは、収量性の低さと作柄が不安定であることが課題となっています。その理由としては、以下のことが考えられます。フツウソバは、異形花型自家不和合性に基づく他殖性植物であり、結実するためには、短柱花と長柱花間での受粉が必要となります。両花形間の花粉輸送は、ミツバチなどの訪花昆虫が担当しています。近年の破壊的な気候変動の影響で、昆虫等の活動が著しく低下しています。そのため、結実を訪花昆虫に依存しているフツウソバでは、作柄が極めて不安定になってしまいます。私たちは、この課題を解決するために、S-del1 を自殖性の供与親として、フツウソバの生殖様式を他殖性から自殖性に改変しました。それにより、夏ソバの普及品種「キタワセソバ」の遺伝的背景をもつ系統NAK-1の開発に成功しました。 従来のフツウソバ品種は、1株に50粒程度の種子が着粒します。それに対し、コムギでは300粒程度、イネでは1,000粒程度の種子が着粒します。私たちが開発したNAK-1は、300粒程度の種子が着粒します。したがって、コムギの収量性に匹敵する、従来のフツウソバの6倍程度の収量性を示す系統の開発に成功したと言えます。さらに NAK-1は、訪花昆虫の活動が低い状態でも1株に多くの種子が着粒するため、気候変動対応型の特性を備えていることも確認できました(図 3)。NAK-1は「キタワセソバ」の遺伝的背景をもつことから、春播き栽培で高い収量性が期待されます。現在、私たちは NAK-1を軸に、消費者・生産者・実需者ニーズに応える健康機能性成分等を含む高付加価値品種の開発に取り組んでいます。
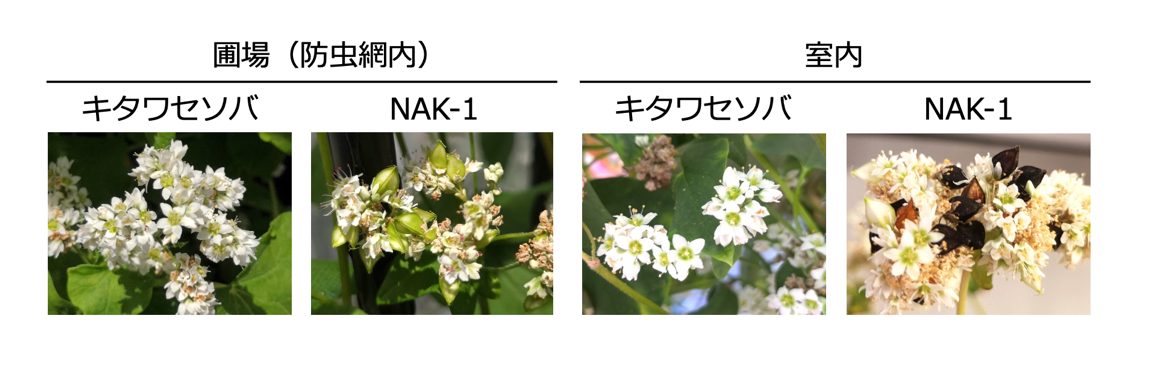
図3 訪花昆虫不在下におけるNAK-1の種子生産能力
訪花昆虫がフツウソバの花に接触が困難な状態(防虫網内や室内栽培)では、従来の他殖性ソバ(キタワセソバ)の結実は認められません。ところが、NAK-1では、たくさんの結実を認めることができます。
教員紹介
相井 城太郎
准教授
学位:博士(農学)
中野 絢菜
助手
学位:学士(応用生命科学)
先生からのメッセージ
みなさん、いつもとちがう「そば」、「新奇な食感・食味をもつそば」を食べてみたくはありませんか?私たちは、圃場と実験室を行き来し、「新奇な食感・食味をもつそば」の開発を目指し頑張っています。植物体に触れ、そのものがもつ生物学的情報を読み解き、得た情報を利・活用することで新しい植物を創る、こんな「わくわく」「ドキドキ」する研究を一緒にしてみませんか。
